Construction of smart phosphor with neuromorphic behaviors
Biological synapse is the fundamental unit of neural information processing, which consists of presynaptic and postsynaptic terminals separated by a synaptic cleft (Fig. 1a). This intricate structure facilitates neurotransmitter-mediated communication, inspiring computational models that emulate synaptic plasticity and adaptability for more efficient information processing39,40. PL-based neuromorphic computing offers potential advantages over electrical and optoelectrical systems including high information bandwidth and visualizable results. However, most existing PL-based neuromorphic systems are constructed upon certain photophysical processes26,27,28. Their continuous decay characteristic poses significant challenges in further developments, particularly in terms of parallelism and information security. To better address these challenges, we categorize current PL-based neuromorphic phosphor systems into two types: Decay-type, constructed upon certain photophysics phenomena with synaptic post-stimulation PL signal, leading to passive Write-only operations; Variation-type, exhibit history-dependent PL properties variations in dark states originated from light-induced material reconfigurations, which enables discrete Write and Read operations.
Fig. 1: PL-based Write and Read mechanisms of MHPe@MYE.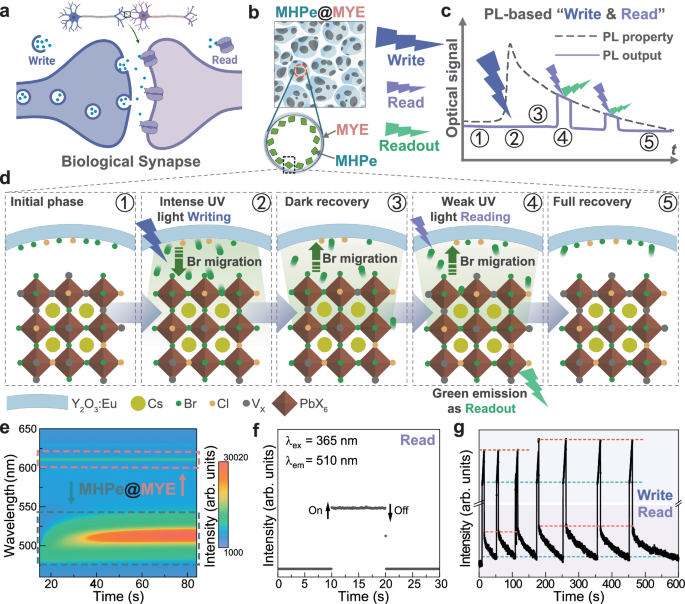
a Schematic of a biological synapse. Created in BioRender. Lao, X. (2025) https://BioRender.com/syuovcp. b Schematic illustration of the MHPe@MYE porous structure. Created in BioRender. Lao, X. (2025) https://BioRender.com/at18ev9. c PL properties variation of MHPe@MYE showing PL properties changes during Write and Read operations, with tagged points (1–5) corresponding to specific PL states. d Schematic illustrations of the microscopic mechanisms at points 1–5 in (c). e Time-resolved PL intensity mapping of MHPe@MYE upon continuous 360 nm laser excitation. The tagged area by blue and pink dash line represents PL signal from MHPe and MYE, respectively. (Color bar: blue to red represents 1000 to 30020 in arb. units). f Inert PL dynamics of MHPe@MYE under Read excitation. g PL dynamics featuring both Read and Write processes.
Such integration of Read operation in PL-based systems fundamentally relies on materials whose light-induced property changes can give rise to PL variations with neuromorphic characteristics, together with a non-destructive probing mechanism to read out the current PL state. The potential advantages and importance of developing Read operation in PL-based neuromorphic systems are discussed in Supplementary Note 1, illustrating that the proposed variation-type may align more closely to the requirements for practical PL-based neuromorphic computing.
To actualize this conceptual framework, we develop MHPe@MYE smart phosphor with porous composite structure (Fig. 1b). The rich interfacial interactions within MHPe@MYE enable light-induced PL properties variations and recovery in dark states, giving rise to dynamics PL features (black dot line) and on-demand Read operations (lavender arrows) as illustratively plotted in Fig. 1c. This adaptive variation is primarily attributed to light-induced halogen migration at the interfaces within MHPe@MYE. Fig. 1d presents a flow diagram of the microscopic mechanisms upon high-energy Write, dark recovery, and low-energy Read, corresponding to the frames tagged 1–5 in Fig. 1c. Starting from interfacial regions containing defective MHPe and Br/Cl-anchored MYE surface (Frame 1), the adsorbed halogen species (Br/Cl) would migrate to MHPe under intense Write light stimulation (Frame 2). It results in altered halogen composition and defect passivation of MHPe, consequently modifying its PL properties. Upon removal of the light stimulus, the PL properties gradually recover in dark states due to reverse migration of halogen species (Frame 3). For implementing Read operation, low-energy probing light is employed, which does not induce further PL variation, thus allowing for non-destructive probing of the current PL state (Frame 4). The system could return to its initial state after long-term resting (Frame 5). This reversible process facilitates effective Write and Read operations, underpinning the neuromorphic functionality of MHPe@MYE for PL-based information processing. Meanwhile, the inert Eu3+ luminescence from MYE host can further serve as a baseline for calibrating the PL readouts.
MHPe@MYE is synthesized following the hard-templated sol-gel procedure37. Aligned with our design, the kinetic PL mapping of MHPe@MYE under continuous 360 nm laser excitation (Fig. 1e) shows characteristic PL variations featuring a red-shifted and drastically enhanced PL intensity at the green region (cycled by blue dot lines), as well as a steady Eu3+ peak at around 613 nm (marked by pink dot lines). The decay process of the PL states can be probed using low-energy Read excitation in non-destructive way (Fig. 1f). Read excitation also reveals the initial PL characteristics featuring a weak peak at 470 nm from defect-rich and Cl-rich MHPe (Supplementary Fig. 2). Using alternating 360 nm laser illumination as Write and weak 365 nm excitation as Read, the PL dynamics of MHPe@MYE are investigated. As shown in Fig. 1g, it shows an enhancing PL intensity upon Write excitations, consistent with the kinetic PL mapping results. Multiple cycles of alternating Write and Read operations reveal consistent recovery endpoints during Read and starting points of subsequent Write processes (marked by green dot lines). Similarly, consistency is observed between orange dot lines marking the end of Write and the start of Read operations. These consistencies confirm equivalent PL states at these critical points, despite different PL intensities during Write and Read conditions. It ensures the foundation for reliable operations in MHPe@MYE artificial synapses.
To clarify the structural underpinnings of the PL variations in MHPe@MYE, a series of material characterizations are carried out. X-ray diffraction (XRD) analysis confirms the good crystallinity and high phase purity of MHPe@MYE. The phase ratios of the composites are calibrated using XRD refinements, showing a majority of MYE and minor MHPe (95.0% v.s. 5.03%, Supplementary Fig. 3a). Transmission electron microscopy (TEM) and scanning electron microscopy (SEM) reveal the uniform porous structures over the samples (Supplementary Fig. 3b). The corresponding energy dispersive spectrometer (EDS) spectra and element mapping also demonstrate homogeneous distribution of all constituent elements (Supplementary Fig. 3c–j), confirming the formation of extensive interfacial regions that are crucial for the light-induced and dark-recoverable PL variations.
Theoretical insight on adaptive photoluminescence variation
The PL variation in MHPe@MYE is primarily driven by light-induced interface-controlled halogen migration, leading to composition variations from defective Cl-rich to Br-rich MHPe. While halogen migration under light irradiation is common in hybrid and mixed I/Br halide perovskite systems, it is rare in all-inorganic Cl/Br systems due to their structural robustness41,42. Comparison studies show no PL variation observed in CsPb(Br/Cl)3 (CPBC) polycrystalline films or nanoparticles (refer to Supplementary Note 2 for details). On the other hand, the halogen migration process in metal-halide perovskites is known to be highly susceptible to surface effects43. Meanwhile, porous materials have demonstrated vast potential in providing interface interactions in luminescent materials38,44. In this context, the porous interface unlocks this otherwise challenging halogen migration in all-inorganic Cl/Br systems, thus prompting a detailed mechanistic investigation.
To this end, DFT simulations are carried out to investigate the role of MHPe@MYE interfaces in the halogen migration process. As demonstrated in Fig. 2a and Supplementary Fig. 5a, the introduction of halogen defects primarily increases the bandgap with increasing Cl content. It does not lead to considerable variation in the band structure of CsPbBr3 (CPB), CPBC, or halogen-defective lattices (denoted as v-CPB and v-CPBC), attributing to the defect tolerance of lead halide perovskites45. Meanwhile, the large bandgap (4.2 eV) makes Y2O3 inert in the PL variation process, and it only provides space to incorporate Eu3+ dopants and 5D0 → 7FJ emission (Supplementary Fig. 5b). Halogen vacancy defects have low formation energies and commonly exist in all-inorganic perovskite crystals46,47. On the other hand, Y2O3’s strong adsorption of Br/Cl can create defect-rich MHPe at the porous channels, as evidenced in previous work37,38. The surface energies (\(\gamma\)) of Br/Cl adsorption on different symmetric sites of Y2O3 surfaces are then compared (Fig. 2b). It shows that values of Br-adsorbed models are always larger than the corresponding Cl counterparts (Fig. 2c), indicative of the comparably unstable Br adsorption. The charge density difference (CDD) diagrams of halogen-adsorbed Y2O3 also demonstrate the stronger charge interactions between Y and Cl rather than Br as compared in Fig. 2d, attributing to the higher electronegativity of Cl (3.0 vs. 2.8 for Br) and smaller ionic radius (1.81 Å vs. 1.96 Å).
Fig. 2: Theoretical insights into the adaptive halogen migration behaviors of MHPe@MYE.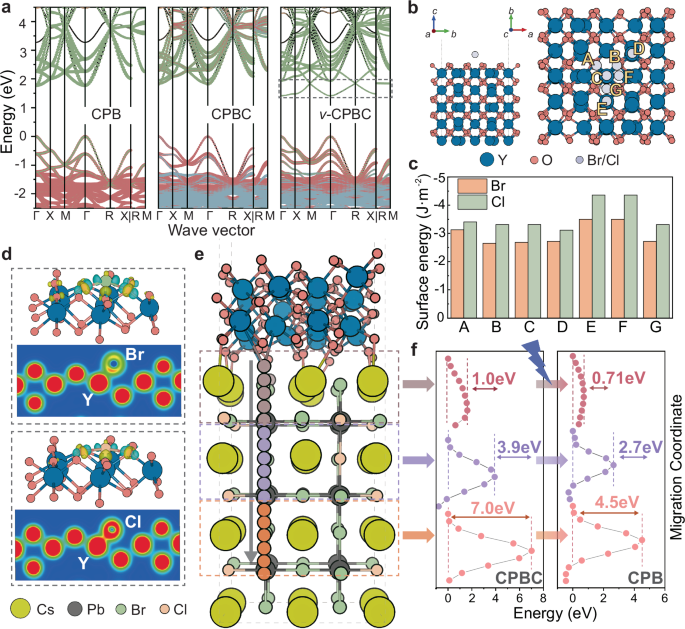
a Electronic band structure of perfect CPB, perfect CPBC, and v-CPBC. The green, red, and blue regions indicate the projected contributions from Pb-, Br-, and Cl-orbitals, respectively. The dot line cycles the emergent shallow defect level. b Illustrative scheme of the adsorption site of Br/Cl on Y2O3 surface and c the corresponding \(\gamma\). d CCD diagram of Br- and Cl-adsorption on Y2O3 surface, where red and blue region indicates electron accumulation and depletion, respectively. e Schematic illustration of Br migration pathway from MYE-surface adsorption and interface to subsurface and inner CPBC (or CPB) lattice. f The NEB energy landscape of the migration processes in CPBC and CPB.
Next, the interface-controlled halogen migration process is investigated from anchored Br/Cl on the MYE surface to the surface site of MHPe, and finally into the deeper MHPe lattice by the nudged elastic band (NEB) method. The model featuring an interface between Y2O3 and CPBC is illustrated in Fig. 2e. First, we examine Br migration into a CPBC lattice and calculated the migration energy barriers at each layer (Fig. 2f). Detaching from the surfacial sites of Y2O3, Br needs to overcome a small energy barrier of 1.0 eV to reach the surface of MHPe. This process mirrors the initial stage of empirical PL variations, where Br can detach from Y2O3 and migrate to the surface of defect-rich MHPe upon illumination. Since surface defects and their passivation are pivotal in determining the quantum efficiency of halide perovskites, this process would lead to enhanced luminescence intensity48,49. Further penetration of Br into CPBC faces substantially higher barriers (3.9 eV and 7.0 eV for subsurface and inner layers, respectively). Meanwhile, as the Br migration process proceeds, the halide composition of MHPe also evolves, potentially altering the migration energy barrier. The model of Br migration from Y2O3 surface into the CPB lattice is then investigated, which ideally coincides with the final stage of empirical PL variations with dominated green light emission from CPB. Compared to the CPBC case, the interface barrier of Br migration decreases to only 0.71 eV, with subsequent energy barriers also decreasing to 2.7 eV and 4.5 eV, respectively. This result marks an accelerated Br migration as the Br composition of MHPe increases, explaining the experimentally observed adaptive PL properties evolution with halogen composition. On the other hand, Cl migration faces higher barriers, consistent with dominant green CPB emission during PL evolution. In contrast, bulk CPB shows a higher Br migration barrier (2.3 eV, Supplementary Fig. 6), highlighting the critical role of MHPe@MYE interfaces in enabling adaptive PL changes with neuromorphic behaviors.
Neuromorphic behaviors in photoluminescence variations
Building upon the theoretically adaptive halogen migration behaviors of MHPe@MYE, the neuromorphic properties of MHPe@MYE are then studied through PL dynamics using programmed light pulses as excitation sources. Fig. 3a illustrates the synaptic PL behavior of MHPe@MYE by monitoring emissions at 510 nm (MHPe) and 613 nm (MYE). A 365 nm LED powered by an arbitrary waveform generator (AWG) setting at 2 Hz is used as excitation sources. The initial stage requires 85 high-energy Write excitation pulses (~161.15 μW·cm−2) to reach the upper threshold (blue line). The subsequent dark recovery process is monitored using low-energy probing pulses (~7.96 μW·cm−2). The lower threshold (lavender line) is reached within time periods equivalent to 72 Read pulses. Notably, subsequent cycles show adaptive responses, with the second round requiring only 36 Write pulses to reach the upper threshold but 90 Read pulses to recover. This indicates MHPe@MYE’s ability to memorize recent excitations, accelerating PL variation while slowing recovery. This effect becomes more pronounced in the third round, with 32 Write and 112 Read pulses, whereas it slightly diminishes after a 1-min interval in the fourth round (51 Write and 89 Read). This behavior mirrors the experience-dependent plasticity in biological synapses, where prior stimulation history influences system responses. In sharp contrast, the Eu3+ emission from MYE exhibits constant binary On/Off states dictated solely by excitation power without neuromorphic features. The coexisting neuromorphic and inert PL behaviors within the MHPe@MYE system serve as an internal reference. To further characterize these neuromorphic properties, detailed investigations into PL variation process are conducted in search of specific biological phenomena and metrics.
Fig. 3: Neuromorphic behaviors in the PL variation process of MHPe@MYE.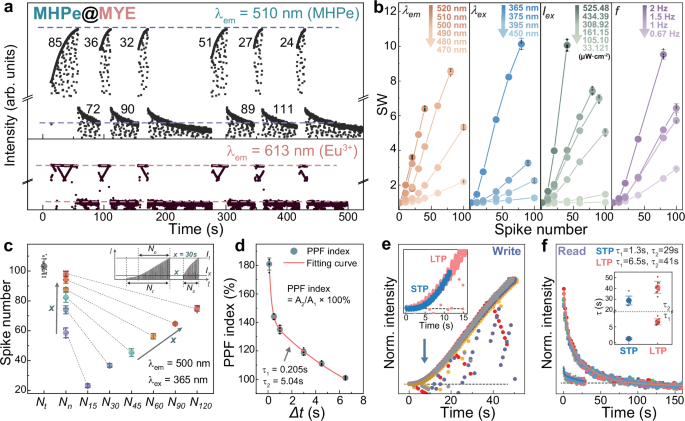
a PL dynamics of MHPe (upper) featuring Write PL variation and Read dark recovery process, and the PL dynamics of Eu3+ in MYE (lower) during the same process using programmed 365 nm LED as excitation light spikes. b Stimuli-dependent synaptic behaviors of MHPe@MYE under different spike parameters of λem, λex, Iex, and f. Error bars are s.d. from four measurements. cx-dependent LTM features of MHPe@MYE, error bars represent s.d. of five measurements. Inset shows the definitions of Nn, Nt, Nx, It, and Ix in the PL dynamics (e.g., x = 30 s). d Fitting curve of PPF indices of MHPe@MYE with s.d. error bars from four measurements. e PL dynamics during formation of STP and LTP. Inset shows the reproducibility of STP (blue) and LTP (red) at the early stages of PL dynamics. f PL dynamics during the decay of STP and LTP, and the corresponding fitting curves. Inset displays s.d. statistical analyses of the fitted time constants across eight measurements.
In biological neurons, synaptic plasticity allows a synapse to adjust its synaptic weight (SW) in response to stimuli. In PL-based artificial systems, SW can be quantified by comparing the PL intensity before and after excitation spikes, and the SW variation trends under different stimulus conditions reflect the stimuli-dependent synaptic behaviors. Our study investigates these behaviors across varying spike parameters specifically based on PL features, including emission wavelength (λem), excitation wavelength (λex), excitation intensity (Iex) and frequency (f). The results are summarized in Fig. 3b, with raw spectral data displayed in Supplementary Figs. 7–10. Taking the λem series as a reference, when monitoring shorter wavelengths (470–490 nm), SW grows rapidly within 20 spikes but saturates at small values, which is due to the red-shifting wavelength at initial stages. The largest SW is observed at 500 nm, with longer wavelengths showing slower evolution and smaller SW. These trends highlight the diverse synaptic behaviors across different λem. Notably, as shown in Supplementary Fig. 7, after 45 s intervals, fewer spikes are required for MHPe@MYE to reach the same SW values, attributed to the history-dependent adaptive behavior of MHPe@MYE. Similarly diverse synaptic evolution trends can be observed in the Iex series. As demonstrated in Supplementary Fig. 9, the SW values and variation speeds decrease monotonically as Iex decreases, whereas the accelerated recovery behavior persists after resting periods. When Iex is further reduced to ~33.12 μW·cm−2, approaching the energy density of Read (~7.96 μW·cm−2), PL variations can no longer be observed. The dependence of SW is similarly studied when varying other parameters of excitation spikes (f, λex), showing that higher SW correlates with shorter λex and higher f. Reproducibility of such stimuli-dependent synaptic behaviors are examined across four measurements with tolerable standard deviation (s.d.). These rich and adjustable neuromorphic behaviors through multiple spiking parameters enhance the information bandwidth and offer flexibility in designing PL-based neuromorphic systems, such as the potential task-adaptive parameters tunning50. Moreover, this ability to sustain neuromorphic characteristics after resting periods is similar to the long-term memory (LTM) phenomena.
LTM behavior represents a sustained synaptic strength modulation over extended periods, driven by mechanisms like receptor activation and Ca²⁺ influx that stabilize experience-dependent memory in biological origins51. As shown in Fig. 3c, we observe analogous LTM behavior in MHPe@MYE from the recovery and recalling processes from a highly potentiated PL states after various resting time (denoted as x). The inset spectrum demonstrates how LTM behaviors are quantified by comparing Nt, Nn, and Nx, as defined in Supplementary Note 3. Statistical analysis of five independent measurements confirms high consistency, as evidenced by the minimal error bars in Fig. 3c (raw data in Supplementary Fig. 11). It demonstrates that a smaller x leads to a higher Ix and fewer Nx required to arouse the same It again, indicating good preservation (smaller Nn) and fast recalling (smaller Nx) of memory after short periods. As x prolongs, Nn and Nx become larger and move close to pristine Nt (indicated by gray arrows in Fig. 3c), mimicking the gradual fading (lager Nn) and hard recalling (larger Nx) of memory after long intervals. When resting intervals exceed a critical duration (~180 s), the system loses its retained state (Supplementary Fig. 12), analogous to biological forgetting processes. The above behaviors replicate the LTM effects by PL manners, indicating the intriguing long-term neuromorphic behaviors of MHPe@MYE.
On the other hand, pair-pulse facilitation (PPF) is a pivotal indicator for determining the short-term functions of neuromorphic behaviors. It occurs when two consecutive identical pre-synaptic stimuli are applied, leading to a progressively enhanced postsynaptic response. Here, the PPF is measured at higher excitation energy (365 nm, ~668.78 μW·cm−2), while λem is set at 500 nm for they exhibit faster adaptation during the early stages. PPF index can be defined as the PL intensity ratios between two paired pulse excitation at time interval \(\Delta t\) (Supplementary Fig. 13). Supplementary Table 1 gives a comprehensive benchmark of PPF behavior in a wide spectrum of synaptic devices. MHPe@MYE exhibits ~181% PPF index measured at \(\Delta t\)= 0.1 s, which is a modest value among existing reports. PPF index typically exhibits a bi-exponential decay relationship against pulse interval (\(\Delta t\)) regardless of the input/output modes14,52,53,54:
$${{{\rm{PPF\; index}}}}={{{{\rm{C}}}}}_{0}+{{{{\rm{C}}}}}_{1}\exp \left(-\frac{\varDelta t}{{\tau }_{1}}\right)+{{{{\rm{C}}}}}_{2}\exp \left(-\frac{\varDelta t}{{\tau }_{2}}\right)$$
(1)
where \({{{{\rm{C}}}}}_{0}\) is a constant, \({{{{\rm{C}}}}}_{1}\) and \({{{{\rm{C}}}}}_{2}\) represent the initial facilitation degrees of the two phases, and \({\tau }_{1}\) and \({\tau }_{2}\) are the characteristic relaxation times for the fast and slow phases, respectively. Supplementary Fig. 13 presents the measured PPF values at different time intervals, and the resultant fitting curve is shown in Fig. 3d. The fitted \({\tau }_{1}\) and \({\tau }_{2}\) values are 0.205 and 5.04 s (adjusted R2 ≈ 0.99), respectively. The two decay channels are indicative of the fast and slow relaxation phases of PPF: the short \({\tau }_{1}\) value enables immediate signal amplification, while the longer \({\tau }_{2}\) value reflects a more gradual change in facilitation for sustained memory over extended periods. This dual-phase relaxation behavior closely mimics the dynamic response of synaptic facilitation, where both fast and slow processes contribute to the complex temporal and adaptive signal processing.
In biological systems, short-term plasticity (STP) and long-term plasticity (LTP) represent fundamental mechanism underlying learning and memory formation. STP functions as a dynamic filter for temporal information processing and working memory, whereas LTP facilitates the consolidation of significant information into long-term memory via sustained synaptic enhancement55. MHPe@MYE can also emulate STP and LTP behaviors in PL-based manner, and the distinct decay processes can be monitored by Read operations. As shown in Fig. 3e, the PL states of MHPe@MYE can be categorized into STP with fewer Write spikes (blue) and LTP with more spikes (red). The variation trends of both LTP and STP demonstrate good reproducibility across eight different measurements. Decay time constants can be acquired from the decay curves (Fig. 3f) of STP and LTP, respectively, by fitting the bi-exponential decay function:
$$y={y}_{0}+{{{{\rm{A}}}}}_{1}\exp \left(-\frac{\varDelta t}{{\tau }_{1}}\right)+{{{{\rm{A}}}}}_{2}\exp \left(-\frac{\varDelta t}{{\tau }_{2}}\right)$$
(2)
The fitted time constants are larger for LTP (\({\tau }_{1}\) = 6.5 s, \({\tau }_{2}\) = 41 s) than STP (\({\tau }_{1}\) = 1.3 s, \({\tau }_{2}\) = 29 s), with tolerable s.d. error margins as shown in the inset of Fig. 3f. This suggests the capability of MHPe@MYE to emulate enduring synaptic modifications by LTP induction, which allows for sustained potentiation over extended periods compared to STP. Such behavior is consistent with the fundamental principles of synaptic plasticity observed in biological systems.
The reproducibility of MHPe@MYE’s neuromorphic properties is further verified through four new batches. The PL properties and structural characteristics of other MHPe@MYE samples demonstrate high batch-to-batch consistency, showing similar porous structure and history-dependent reversible PL properties variations. Supplementary Note 4 presents a detailed comparison based on representative results. It should be emphasized that the PL-based neuromorphic behaviors observed in MHPe@MYE mainly mimic excitatory processes, which capture the essential nature of memory acquisition, reinforcement, and retention56. Biological synapses also exhibit complementary functions like inhibitory behaviors. Achieving bidirectional optical modulation of optical responses remains a technical challenge due to the unidirectional nature of light, and PL-based inhibitory excitation requires further mechanism innovations. Learning from existing optoelectrical devices, recent reports have demonstrated that heterostructural designs can enable both excitatory and inhibitory control of electrical states by optical stimuli14,57,58. Other studies have successfully employed electrical control or combined optical-electrical architectures to realize inhibitory behaviors52,59. These approaches provide valuable insights into further enriching the PL-based neuromorphic functions of smart phosphor, particularly in terms of heterostructure engineering and strategic introduction of secondary physical terminals.
Physical reservoir computing based on smart phosphor
The above results confirm the intriguing PL-based neuromorphic behaviors of MHPe@MYE, characterized by volatile and nonlinear responses to optical stimuli in ultraviolet (UV) and blue regions. These characteristics align well with the fundamental principles of reservoir computing, including nonlinear signal processing and high-dimensional data projection, which facilitates effective feature extraction and transformation of original dataset before downstream classification60,61,62. To evaluate the feasibility of MHPe@MYE as PL-based physical reservoir, we design an experimental setup using Write pulses (λex = 365 nm, ~668.78 μW·cm−2) as inputs and PL intensities under Read excitation (λem = 500 nm, read by ~7.96 μW·cm−2) as outputs, respectively. The custom measurement setup is depicted in Supplementary Note 5. The input signal encoded 4-bit binary sequences (0000 to 1111) as periodic waveforms controlled by AWG (1 Hz), with 1 represented by an active cycle and 0 by absence. An example waveform for the sequence 1011 is shown in Fig. 4a. The measured Read PL dynamics are shown in Fig. 4b, where all 16 Iem-t curves (from 0000 to 1111) exhibit distinct PL dynamics features with notable reproducibility (Supplementary Fig. 15). The results demonstrate that the PL-based MHPe@MYE physical reservoir exhibits stable fading memory characteristics, where the output PL state depends critically on both the number of excitation pulses and their temporal distributions. This can be exemplified by sequences 1100 and 0110 (Fig. 4c), though both contain two pulses, their distinct timing sequence induces different PL trajectories. Similarly, sequences with varying pulse counts (1100, 1110, and 1111) also generate distinct PL variations. This nonlinear weighting of excitation spikes enables MHPe@MYE reservoir to map distinct 4-bit sequence to characteristic PL responses. Furthermore, sampling points (SMPs) are discrete time instants at which readouts are sampled from the PL dynamics of MHPe@MYE reservoir, thereby encoding temporal PL responses into static features. As tagged in Fig. 4b, two SMPs are manually assigned based on inter-class separation among 16 PL dynamics. While PL readouts at individual SMP show high reproducibility with minimal s.d. errors among eight experiments, neither could fully differentiate all 4-bit codes due to signal overlaps in the one-dimensional feature space (Fig. 4d). By contrast, plotting paired SMP readouts can transfer the time-resolved waveforms into a two-dimensional (2D) feature space of PL intensity map (Fig. 4e), where all 16 binary patterns become clearly and reproducibly distinguishable.
Fig. 4: Potential application of MHPe@MYE as all-optical physical reservoir.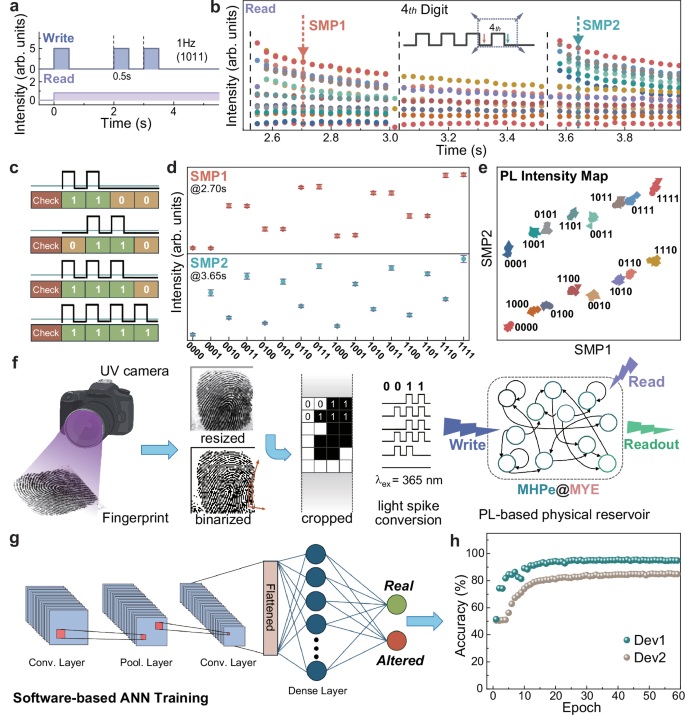
a Schematic representation of 4-bit binary code 1011 encoding through Write and Read light pulse waveforms. b Characteristic Read PL dynamics of MHPe@MYE of 16 binary sequences, with SMP locations indicated in the inset. c Representations of 1100, 0110, 1110, and 1111 Write waveforms used as binary sequences. d Statistical quantification of PL readout distributions at SMP1 and SMP2 of all 16 binary sequences, demonstrating code-specific PL signatures with small s.d. error margins from eight measurements. e 2D feature space projection of dual-feature PL readouts based on eight measurements, showing effective separation of all 4-bit sequence. f Schematic illustrations of SOCOFing fingerprint data acquisition and preprocessing from UV camera imaging, and light pulse conversion to full-PL-based information processing through the MHPe@MYE physical reservoir. Scheme was created in BioRender. Lao, X. (2025) https://BioRender.com/qgctjin. g Software-based offline ANN training of the processed results for distinguishing authentic and counterfeit fingerprints. h Training records of Dev1 and Dev2.
To validate the practical utility of MHPe@MYE as physical reservoir, it is first applied to process MNIST dataset for handwritten digit recognition63. The MNIST images are preprocessed into stripes of 4-bit binary codes, with detailed methodology outlined in Supplementary Note 6. To compare different SMP designs, three virtual devices with varying abilities to distinguish 4-bit binary codes are constructed. Dev1 employs two SMPs for dual-feature sampling, ensuring better input code identification according to the well-separated features in Fig. 4e. In contrast, Dev2 and Dev3 use only SMP1 or SMP2 as the readout, respectively. The experimental PL readouts are extracted from Fig. 4d with different SMP configurations, which are then fed into a simple artificial neural network (ANN) for offline software-based training to evaluate the MNIST recognition accuracy after PL-based physical reservoir computing. As plotted in Supplementary Fig. 17, the training results show that Dev1 can achieve classification accuracy of 92% within 20 epochs, while Dev2 and Dev3 reach only 90% and 87%, respectively. This result highlights that the dual-feature method offers better performance in PL-based physical reservoir computing.
To further validate the capability of PL-based reservoir computing in more complex application scenarios, their potential in UV light-based fingerprint recognition is then explored. While optoelectronic devices are typically employed for processing UV light inputs, our study shows that MHPe@MYE can effectively replicate this process through PL-based manner64,65,66. The workflow of proposed MHPe@MYE physical reservoir for SOCOFing fingerprint authentication is illustrated in Fig. 4f and described in Supplementary Note 7, as modified from previous reports based on optoelectronic devices64. After optimizing the binarization and resizing parameters of SOCOFing images in Supplementary Figs. 18–20, we select the adaptive thresholding binarization and 64 × 64 resizing pixels due to their good performance in balancing training costs and preservation of features. A similar approach is then used to transform the resized images into 4-bit binary codes (Supplementary Fig. 21a, b), which further modulate the input waveform to MHPe@MYE PL-based physical reservoir. Experimental PL dynamics maps are again extracted as the readouts, and fed into ANN to evaluate the recognition accuracy (Fig. 4g). The training results are plotted in Fig. 4h, showing that the dual-feature method (Dev1) leads to much higher accuracies throughout the training process, and finally reaches 94.4% after 60 epochs. The results are comparable to optoelectronic neuromorphic devices despite the different information processing modes and algorithms15,16,64,67. In sharp contrast, the result of Dev2 (employing SMP2 only) attains markedly lower accuracy of 84.2%. The disparity between dual- and single-SMP methods is consistently observed across different resizing strategies as compared in Supplementary Fig. 21c–e. Supplementary Fig. 21f summarizes the accuracy and loss of SOCOFing recognition training process at different resize methods.
The above PL-based physical reservoir concept can be broadly extended to other smart phosphor systems, such as the decay-type PersL phosphors, as exemplified in Supplementary Note 8. Nevertheless, the absence of Read mechanism impedes the effective retrieval of information due to internal and external interferences. Comparative studies reveal that decay-type systems experience notably increased computational costs and decreased accuracy resulting from wrong readouts and feature-loss events, particularly with decay signal overlaps from adjacent units (details in Supplementary Note 9). This marks the importance of achieving distinct Read operations for future developments of PL-based neuromorphic computing.